
東北大、海馬などの神経幹細胞の分裂に脂肪酸結合タンパク質の関与を解明 [2012/05/15]
東北大学(東北大)は、成体マウスの海馬歯状回における「神経幹細胞」に「脂肪酸結合タンパク質(Fabp:fatty acid-binding protein)」の「Fabp7」及び「Fabp5」が存在していること、このタンパク質をどちらか一方でも生まれつき欠損したマウスでは神経幹細胞の分裂が減少するが、両タンパク質を同時に失ったマウスでは、神経幹細胞の分裂は減るものの、逆に新生神経細胞の生存が向上することを明らかにしたと発表。さらに、この神経幹細胞の分裂と生存が海馬の前後軸に沿って異なる分布を示すこと、Fabp7及びFabp5を欠損したマウスではこの前後軸における分布にも異常があることも見出したことも発表した。
成果は、東北大大学院医学系研究科の大隅典子教授、松股美穂研究員(現・理化学研究所)らのグループと、山口大学医学部の大和田祐二教授、理化学研究所の吉川武男チームリーダーらとの共同研究によるもの。研究の詳細な内容は、米科学誌「Stem Cells」Web版6月号にまもなく掲載の予定だ。
ヒトの脳では膨大な数の神経細胞がそれぞれ別の神経細胞と無数に結合し、回路網を作って恒常的に情報をやり取りしている。ヒトを含む高等動物の脳は、かつては胎生期に完成すると考えられ、脳を構成する神経細胞の数は生後は減少するのみと信じられていた。
これらの神経細胞の大部分は出生前に作られるのは事実だが、近年の研究により、現在では「側脳室」に面した「脳室下帯」及び「海馬歯状回」のような脳の一部の限られた領域には、生後でも神経細胞を新たに作り出すことのできる種のような神経幹細胞が存在し、この細胞が分裂を繰り返すことにより絶えず新しい神経細胞が作られていることがわかってきた。
またこのような神経新生の異常が、統合失調症や心的外傷後ストレス障害(PTSD)と関係することが報告されており、神経新生のメカニズムについて大きく着目されている状況だ。
哺乳類では、脳室下帯と海馬歯状回の「顆粒細胞下帯」が、そのような神経新生が起こる場所として知られている(画像1)。特に海馬歯状回の新生顆粒細胞は、海馬の主な脳機能である記憶や学習行動に関係していることがすでにわかっており、研究グループではこれまでも海馬歯状回をターゲットとした研究を進めてきた。
画像1は、マウスにおける、成体においても神経幹細胞が存在し神経新生が起こり続ける2つの脳領域の模式図だ。左側が前方、右側が後方で、マウスは左を向いている。
図中、黒の円で囲った赤い丸印は神経幹細胞で、前方の側脳室脳室下帯にいる神経幹細胞から分裂した細胞は分化しながら前方へ移動し(画像中の矢印)、最先端に存在する嗅球という組織で抑制性の神経細胞へと最終分化する。
一方、後方にある海馬歯状回顆粒細胞下帯にいる神経幹細胞は、興奮性の顆粒細胞を産生し、歯状回の中心的役割を担う顆粒細胞層へ新規神経細胞を提供し続ける仕組みだ。
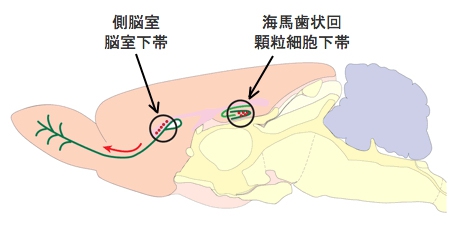
画像1。生後神経新生の起こる脳領域
脂肪細胞を別にすれば、脳は体内で最も「脂肪酸」の含有量が多い組織の1つだ。脂肪酸は通常、脳の細胞の細胞膜の中につなぎとめられているが、神経伝達などの刺激によって細胞膜から遊離したり、あるいは食べ物から摂取されて血流に乗り脳まで到達した場合、脂肪酸結合タンパク質(Fabp)と結合することにより細胞質内において安定化する。
今回、研究グループはこの脂肪酸結合タンパク質の内、脳において多量に存在するFabp7とFabp5に着目し、これらの遺伝子を欠損した成体マウスでの解析を行った。
なお、Fabp7は脳発生初期の未分化な神経幹細胞の中に多量にあり、分化したニューロンにはほとんど見当たらないことから、Fabp7は神経新生の過程において、未分化な神経幹細胞の増殖あるいは分化に関わっているのではと考えられている。Fabp5はFabp7の類縁だが、これまで脳における機能はほとんどわかっていなかった。
今回の研究では、まず野生型の成体マウスの海馬歯状回でのFabp7及びFabp5の発現を詳細に解析。Fabp7及びFabp5は、神経幹細胞の最も未分化な時期から共に脳内に豊富に発現しているが、幹細胞が分裂・分化していく過程において徐々に発現が低下していく。
その後、Fabp7の発現は神経細胞へ分化した時点でほぼ消失するが、Fabp5の発現は続き、よく似たタンパク質でありながら両者は役割の違いがあることが推測された(画像2)。
画像2は、神経幹細胞の分化に伴った、海馬歯状回顆粒細胞層下帯におけるFabp7及びFabp5の発現の変化を撮影したものである。左側(A-G)の緑のシグナルがFabp7の、右側(H-N)の緑のシグナルがFabp5の局在を示す。
(A,H)Fabp7(A)及びFabp5(H)はどちらも海馬全域で発現しているが、特に歯状回顆粒細胞下帯(矢印)において強い発現が見られる。(B-G)Fabp7は、神経幹細胞を示すGFAP(B)、Nestin(C)、SOX2(D)と共局在する細胞が多く、分化していくと発現が低下していく(神経前駆細胞:DCX(E)、神経細胞:NeuN(F))。
上の画像での様子をグラフ化したものが(G)と(N)だ。(B-E)の矢尻は、Fabp7と各種遺伝子を共発現している細胞を示す。(I-N)一方、Fabp5は神経幹細胞を示すGFAP(I)、Nestin(J)、SOX2(K)との共局在も見られるが、神経前駆細胞を示すDCX(L)と共局在する割合が最も高い。さらに神経細胞(M)に分化してからも発現が見られる。なお、(I-M)の矢尻は、Fabp5と各種遺伝子を共発現する細胞を示す。
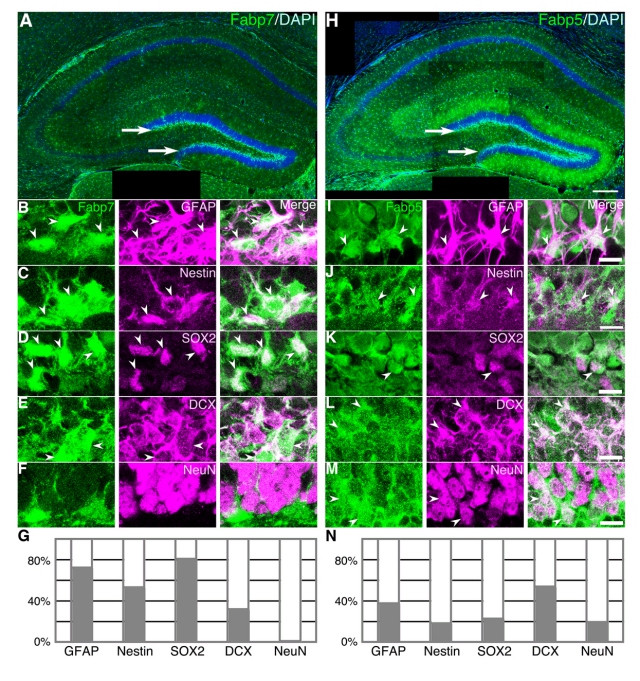
画像2。神経幹細胞の分化に伴った、Fabp7及びFabp5の発現の変化
次にFabp7、Fabp5の欠損マウス(Fabp7KO(7KO),Fabp5KO(5KO))を掛け合わせることにより、両方のFabpタンパク質を欠損したマウス(Fabp7/Fabp5KO(7/5KO))を作成。いずれの欠損マウスも見かけ上異常はなく、脳も一見したところ野生型との違いはない(画像3)。 画像3。野生型マウス(WT)とFabp7KOマウス、Fabp5KOマウス、Fabp7/Fabp5KOマウス。3種類のFabp欠損マウスは野生型と外見的な差異はない
画像3。野生型マウス(WT)とFabp7KOマウス、Fabp5KOマウス、Fabp7/Fabp5KOマウス。3種類のFabp欠損マウスは野生型と外見的な差異はない
しかし、海馬歯状回の神経幹細胞及び分化過程にある細胞を詳細に調べると、欠損マウスではこれらの新生神経細胞の数や突起が少なくなっていることがわかった(画像4)。
そこで、細胞の分裂を標識する化学物質「BrdU」を用いて、Fabp欠損マウスで神経新生が変化しているかどうかが調べられた。投与1日後のBrdU陽性細胞を比較したところ、Fabp7KO及びFabp5KOマウスでは野生型に比べてそれぞれ15%減少しており、Fabp7/Fabp5KOマウスでは30%の減少が見られた具合だ(画像5)。Fabp7/Fabp5KOマウスでは、Fabp7欠損とFabp5欠損の効果が加算されたものと思われる。
なお画像4は、Fabp欠損マウスにおける神経幹細胞、神経前駆細胞の様子だ。野生型マウス(WT:1列目)と比較して、Fabp欠損マウスでは神経幹細胞を示すGFAP(A)、SOX2(B)、神経前駆細胞を示すDCX(C, D)のシグナルが減少している(7KO:2列目、5KO:3列目、7/5KO:4列目)。
画像5は、野生型マウスに比べて、Fabp7KOマウス、Fabp5KOマウスでは15%、Fabp7/Fabp5KOマウスでは30%、BrdU陽性細胞数が減少していた。図中のアスタリスク(*)は、統計学的に有意な差があったことを示す。
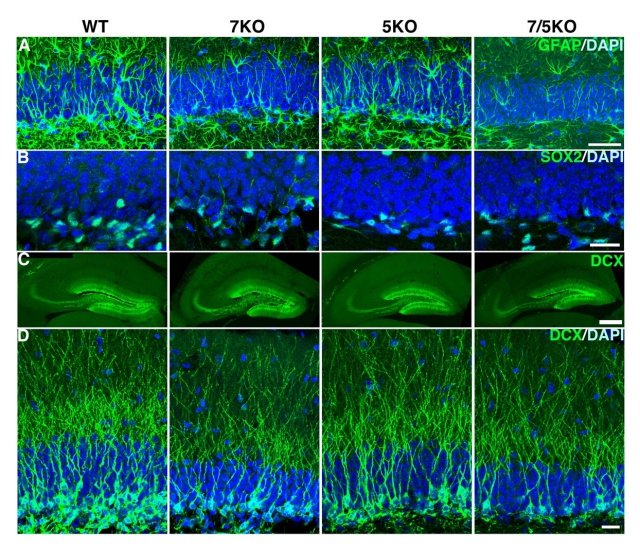
画像4。Fabp欠損マウスにおける神経幹細胞、神経前駆細胞の様子
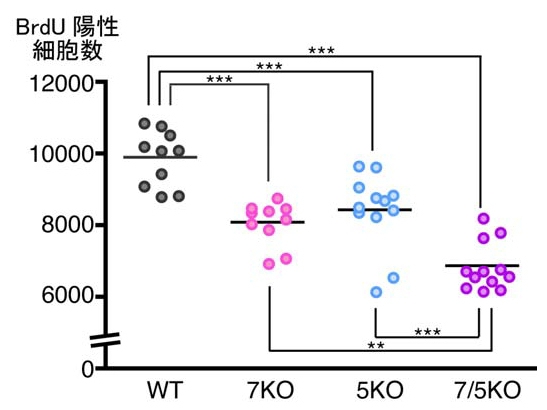
画像5。BrdU投与1日後におけるBrdU陽性細胞数
さらに、投与1カ月後でのBrdU陽性細胞を追跡し、分裂した細胞の生存が調べられたところ、BrdU陽性細胞数は野生型とFabp7KO、Fabp5KOの間では違いがなく、Fabp7/Fabp5KOでのみ増加していた(画像6)。Fabp7とFabp5は細胞の生存に関しては同じ働きをしており、両方がなくなった時に初めて影響が出たと推測される。
画像6は、BrdU投与1カ月後におけるBrdU陽性細胞数。野生型マウス、Fabp7KOマウス、Fabp5KOマウス間ではBrdU陽性細胞数に違いは見られないが、Fabp7/Fabp5KOマウスでは野生型マウスと比較してBrdU陽性細胞数が25%増加していた。
以上のことから、Fabp7とFabp5はどちらも細胞の分裂と生存を適切に制御するように働いていることがわかったのである。
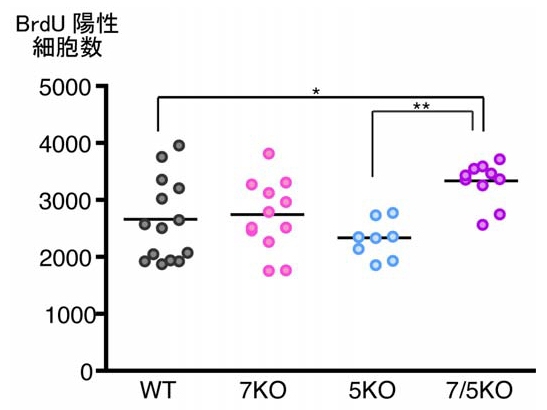
画像6。BrdU投与1カ月後におけるBrdU陽性細胞数
さらに、神経幹細胞の分裂と生存に対する脳の前後軸に沿った分布を調べた。その結果、神経幹細胞の分裂には前後軸に対して前側と後ろ側に2つのピークがあるのに対し、新生神経細胞の生存に関しては前側にのみピークがあること判明(画像7)。
画像7は、BrdU投与1日後及び1カ月後における、BrdU陽性細胞の前後軸に沿った分布BrdU投与1日後(A)及び1カ月後(B)におけるBrdU陽性細胞数を、前後軸に沿った部位別にプロットしたもの(黒:WT、赤:7KO、青:5KO、紫:7/5KO)。
左側がマウス前方、右側が後方にあたる。1日後では、前方と後方の2ヶ所に増殖のピークが見られた(A)。一方、1カ月後のグラフでは、ピークは前方側のみ。アスタリスク(*)は、4種のマウス(WT、7KO、5KO、7/5KO)間で統計学的に有意な違いがあったことを示している。
このことは、前後軸の後ろ側で分裂した細胞が、前側で分裂した細胞に比べて、その後生存しにくいことを示す。Fabpの欠損マウスでは3つの群ともに、この前後軸での分裂や生存の分布にも異常が見られた。
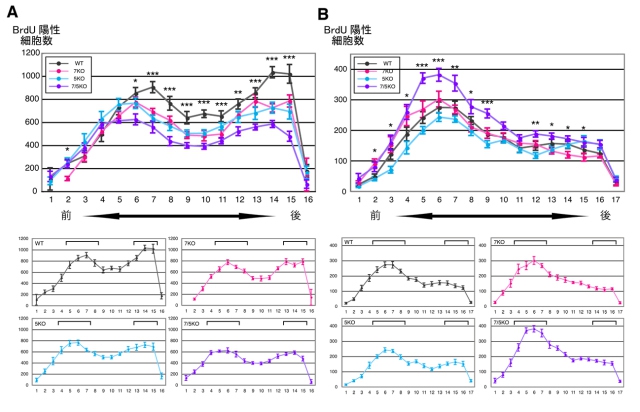
画像7。BrdU投与1日後及び1カ月後における、BrdU陽性細胞の前後軸に沿った分布
生後、ほかの脳部位では起こらない神経新生が海馬で起こることには、海馬依存的な記憶や学習に関係することが知られている。Fabpは、この新しい神経細胞を生み続けていくために、おそらく脂肪酸の細胞内分布などを介して、神経幹細胞の分裂や生存を適切に調節する役割を担っているものと考えられるという。
また、神経幹細胞の分裂で生じた新たな神経細胞が正常に働くためには神経回路にこの新生神経細胞が正しく組み込まれることが必要だが、Fabp欠損マウスでは、今回見出されたように分裂と生存の乱れがあるため神経回路への組み込みに異常がある可能性があり、その回路に依存する海馬の脳機能への影響も示唆されている。
動物モデルにおいては神経新生の低下と精神疾患様の行動異常との間に相関性があることから、今後、栄養素である脂肪酸を用いた適切な介入を行うことにより、例えば心的外傷後ストレス障害(PTSD)などの予防や治療に繋がることも期待される。



