
2013年12月19日
独立行政法人理化学研究所
100年ぶりに脳の主要な記憶神経回路の定説を書き換え
-海馬に新たな記憶神経回路を発見、記憶形成の謎解明へ大きく前進-
ポイント
海馬のCA2領域を多角的かつ正確に同定
海馬で新しいトライシナプス性の記憶神経回路を発見、古典的定義を覆す
神経系変性疾患や精神神経疾患メカニズムの解明に貢献
要旨
理化学研究所(理研、野依良治理事長)は、マウスを使い、脳の記憶形成の中枢である海馬[1]の部位で最も解明が遅れていた領域「CA2[1]」を多角的な手法を使い正確に同定しました。さらに、CA2を介した新しいトライシナプス性[2]の記憶神経回路を発見し、逆に、存在すると主張されてきた回路が、実は存在していないということも証明しました。これは、理研脳科学総合研究センター(BSI、利根川進センター長)RIKEN-MIT神経回路遺伝学研究センター(CNCG)利根川研究室の小原圭吾リサーチサイエンティスト、ミケレ・ピグナテーリ博士研究員、アレックス・リヴェスト博士研究員、利根川進センター長(米国マサチューセッツ工科大学、CNCGディレクター)などが、理研バイオリソースセンターの小幡裕一センター長らと行った共同研究の成果です。
海馬の神経回路については、1911年に神経解剖学者のラモニ・カハールらによって「トライシナプス性の記憶神経回路」が初めて発見されました。その後、1934年に神経生理学者のロレンテ・デ・ノによって海馬が3つの領域(CA1[1]、CA2、CA3[1])からなることが発見され、定義されました。しかし、CA2は他の領域に比べて狭いため、当時の実験技術や装置ではCA2の正確な領域と記憶神経回路を決定することは困難で、その後も生物学的に正しいかどうかは検証されず、現在の教科書でもこの“古典的”な定義に基づいて解説されています。
研究グループは、免疫組織染色法、樹状突起標識法、軸索標識法、および電気生理学などを用いた多角的かつ詳細な観察によって、CA2領域を正確に同定することに成功しました。次に、領域特異的な遺伝子組み換えマウス、ウイルス標識法、光遺伝学、海馬急性スライス標本などを複合的に用いて、海馬の一部位の歯状回[1]が直接シナプスを介してCA2に入力していることを発見しました。この発見は、これまでの「歯状回はCA2に入力しない」という定説を覆すものです。また、CA2はCA1の深い細胞層の興奮性細胞群に優先的に入力していることも分かりました。これらから、カハールらが発見したトライシナプス性の記憶神経回路に加え、新しいトライシナプス性の記憶神経回路があることが分りました。さらに研究グループは、2010年に他の研究グループが発表し、海馬の研究に多大な影響を及ぼした、嗅内皮質3層からCA2への直接の入力は、実は存在しないことも証明しました。
脳の記憶のメカニズムには未解明な部分が多いのが現実です。解明を進めるためには、脳の正確な「地図」が欠かせません。今回の発見は、新技術と既存技術を複合的に組み合わせたアプローチによる成果です。今後、こうした研究アプローチによって、正確でより完成度の高い脳の地図が簡単に作られ、記憶の謎や神経系変性疾患・精神神経疾患のメカニズム解明が進展していくと期待できます。
本研究成果は、英国の科学雑誌『Nature Neuroscience』のオンライン版(12月15日付け)に掲載されました。
背景
私たちや私たちの子孫が、日常社会生活を営んでいくうえで、記憶はなくてはならない脳の重要な機能の1つです。しかし、現在まで、記憶の形成、貯蔵、想起、忘却がどのようにして起きているのかは、ほとんど解明されていません。このため、記憶のメカニズムは生命科学に残された大きな謎の1つです。これまでの臨床例やサルやマウスを用いた実験などから、記憶の形成と想起には、感覚野や大脳連合野などの皮質領域と海馬をつなぐ中継地である嗅内皮質[3]と、海馬の神経回路での正確な情報処理が必要だと考えられています。
1911年に、スペインの神経解剖学者ラモニ・カハール(1906年ノーベル医学生理学賞受賞、図1)らによって海馬のトライシナプス性の記憶神経回路が初めて発見されました。この神経回路は、「嗅内皮質→歯状回(DG)→CA3→CA1」から構成され、3つのシナプスを介することから「トライシナプス性の記憶神経回路」と名付けられました。現在までのおよそ100年間、トライシナプス性の記憶神経回路は、多くの実験研究者、理論研究者によって精力的に研究され、最も有名で重要な記憶神経回路の1つとされています。
一方、海馬は、3つの領域(CA1、CA2、CA3)からなることが1934年にスペインの神経生理学者ロレンテ・デ・ノ(図1)によって発見され、定義されました。しかし、CA2は他の領域に比べて狭いため、当時の実験技術や装置ではCA2の正確な領域と記憶神経回路を決定することは困難でした。また、現在に至るまで、CA2の古典的な定義が、生物学的に正しいのかどうか調べられたことはありません。 最近になり、インシチューハイブリダイゼーション法[4]を用い、CA2とは異なる複数のサンプルをコンピュータ上で合成し擬似的に表示するという間接的な実験結果から、CA2の新しい領域を提唱し、定義する論文も報告されましたが、CA2の新しい領域と古典的な定義の関係性について正確に検証した論文はありません。そのため、現在のほとんど全ての学術教科書のCA2に関する記述は、古典的な定義に基づいています。さらに、その古典的な定義から、「CA2は歯状回からの入力を受けない」とされていました。
そこで研究グループは、マウスを使ってCA2領域の正確な同定に挑みました。
研究手法と成果
研究グループは、CA2領域を正確に同定するために、①免疫組織染色法によるCA2のマーカータンパク質の同時染色(分子生物学的手法)、② ディオリスティック染色法(細胞生物学的手法)、③BDAトレーサー軸索標識法(解剖学的手法)、④パッチクランプ法(電気生理学的手法)の4種類の多角的なアプローチによる実験を行いました。実験の結果、CA2の目印となる主なマーカータンパク質3種類が、共通の領域で強く発現しました(図2)。次に、樹状突起標識法で神経細胞の主要な樹状突起に見られる複雑なスパイン(シナプスからの入力を受けるトゲ状の隆起)の有無、軸索標識法で視床下部の乳頭体上核からの入力の有無、電気生理学的手法で細胞特性などを多角的、詳細に調べました。その結果、CA2領域を正確に同定することに成功しました。
続いて、①免疫組織染色法、②ウイルス標識法、③新規DG特異的遺伝子組換えCreマウス[5]、④新規CA2特異的Creノックインマウス[5]、⑤光遺伝学(色素タンパク質・チャネルロドプシン2を使った実験)、⑥海馬急性スライス標本を複合的に用いて、歯状回のCA2への入力を調べました。その結果、これまで何十年間も信じられていた「歯状回はCA2に入力しない」という定説を覆し、「歯状回が直接シナプスを介してCA2に入力している」ことを発見しました(図3)。また、CA2は、CA1の深い細胞層の興奮性細胞群に優先的に入力していることが明らかになりました。以上のことから、海馬において、従来型のトライシナプス性の記憶神経回路「嗅内皮質→DG→CA3→CA1」に加えて、新しいトライシナプス性の記憶神経回路「嗅内皮質→DG→CA2→CA1deep」を発見しました。従来型のトライシナプス性の記憶神経回路は海馬内で主にラメラ断面[6]に沿って情報伝達しているのに対して、新しいトライシナプス性の記憶神経回路は複数のラメラ断面を縦断して情報伝達していることも明らかになりました。
また、嗅内皮質の3層細胞特異的な遺伝子組換えCreマウスと光遺伝学を用いた実験から、嗅内皮質3層細胞がCA1に入力し、CA2には入力しないことも明らかになりました。これも従来の知見を覆した発見です。
今後の期待
記憶に関わる脳内の神経活動や、さまざまな神経系変性疾患、あるいは精神神経疾患のメカニズムの謎を解明していく上で、脳神経回路の正確な地図は欠かせません。
現在、脳の正確な地図は部分的にしかでき上がっていませんが、脳の地図の完成を目指して、大型の国家プロジェクトが欧州、米国、日本で動き始めました。今回の研究成果は、新技術や既存技術を複合的に用いたアプローチによるものです。このアプローチを用いて、あるいは改良することで、正確でより完成された脳の地図が効率的に作られ、記憶の謎や神経系変性疾患・精神神経疾患のメカニズムの謎の解明がさらに進展していくと期待できます。
原論文情報
Keigo Kohara, Michele Pignatelli, Alexander J Rivest, Hae-Yoon Jung, Takashi Kitamura,Junghyup Suh, Dominic Frank, Koichiro Kajikawa, Nathan Mise, Yuichi Obata, Ian R Wickersham & Susumu Tonegawa.“Cell type–specific genetic and optogenetic tools reveal hippocampal CA2 circuits”Nature Neuroscience,doi:10.1038/nn. 3614
掲載された論文 http://www.nature.com/neuro/journal/vaop/ncurrent/full/nn.3614.html
発表者
独立行政法人理化学研究所
脳科学総合研究センター 理研-MIT神経回路遺伝学研究センター
リサーチサイエンティスト 小原 圭吾 (こはら けいご)
(元日本学術振興会海外特別研究員)
keigo [at] mit.edu
脳科学総合研究センター
センター長 利根川 進 (とねがわ すすむ)
(RIKEN-MIT神経回路遺伝学研究センター 教授)
tonegawa [at] mit.edu
補足説明
海馬、CA2、CA1、CA3、歯状回
海馬は、大脳辺縁系の一部でヒトでは大脳側頭葉の内下部にある。長期記憶の形成や空間学習に重要な役割を果たしている。湾曲した細長い構造がタツノオトシゴ(ラテン語:hippocampus - 英語:sea horse)に似ていることから海馬と名付けられた。特徴的な層構造をした細長い形をしており、CA1、CA2、CA3、歯状回の各部位からなる。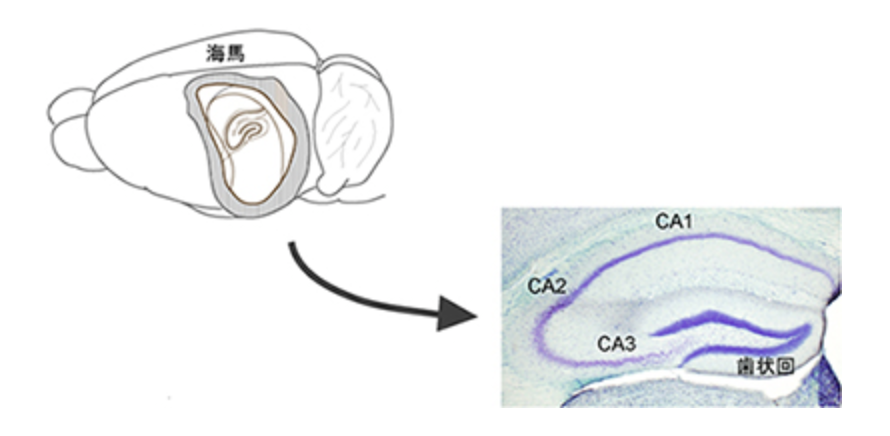
トライシナプス性
異なる3つのシナプスを介することをトライシナプスと呼ぶ。海馬においては、「嗅内皮質→歯状回(DG)→CA3→CA1」から構成される3つのシナプスを介し、記憶の形成と想起に必要な主要な神経回路。
嗅内皮質
空間情報や物体情報の情報処理を行う皮質領域。ここで情報処理された後に、海馬に情報が伝達される。
インシチューハイブリッドダイゼーション法
生体組織の遺伝子発現を調べる実験技術。メッセンジャーRNAに特異的に結合する標識プローブを用いて組織を染色し、いろいろな遺伝子の発現領域、部位を調べることができる。
新規DG特異的遺伝子組換えCreマウス、新規CA2特異的Creノックインマウス
歯状回(DG)特異的な遺伝子Dock10のプロモーター領域の下流にCre遺伝子を挿入した改変遺伝子を染色体に組み込んだ組み換えマウスで、理研バイオリソースセンターと共同で開発した。DG領域のみCre遺伝子が発現する。
CA2特異的な遺伝子Map3K15の遺伝子部位の最後尾にCre遺伝子を挿入したノックインマウス。CA2領域のみCre遺伝子が発現する。
ラメラ断面
補足説明1の図のように、海馬は細長く湾曲したロールケーキのような形状をしている。ロールケーキのどこを切っても同じ切断面がみられるように、海馬も長軸方向に直角に切れば、どこを切ってもDG、CA3、CA2、CA1を含む同じような切断面が見られる。その切断面のことを海馬のラメラ断面と呼ぶ。

図1 ラモニ・カハール(左)とロレンテ・デ・ノ(右)
写真提供:米国国立医学図書館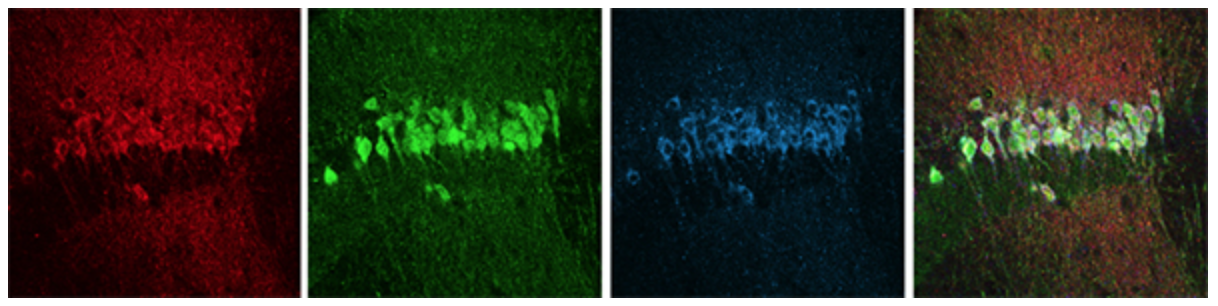
図2 CA2領域のマーカータンパク質の同時染色
CA2領域の目印となるマーカータンパク質を免疫組織染色法により同時染色した。赤色はRGS14タンパク質、緑色は PCP4タンパク質、青色はSTEPタンパク質を示す。右端は3つの画像を重ね合わせたもので、CA2領域を示す。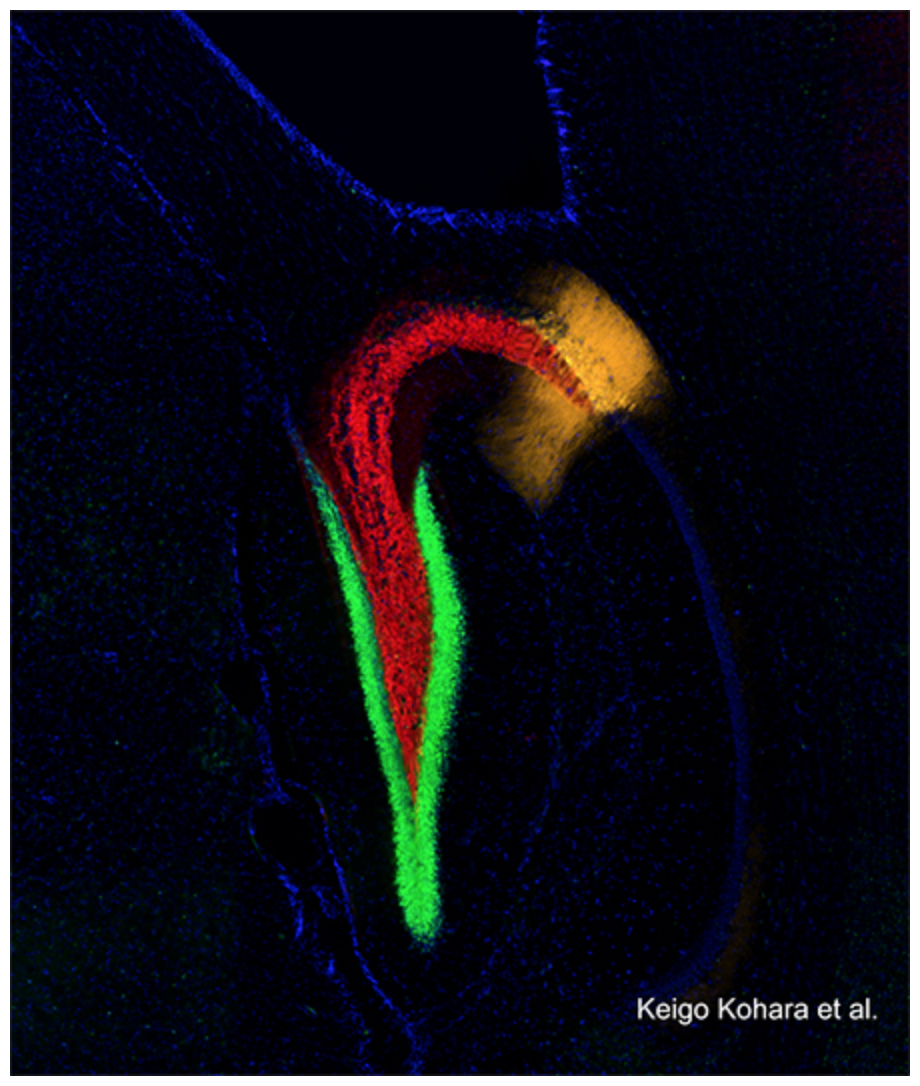
Keigo Kohara et al.
図3 海馬における歯状回(DG)からCA2への新しい記憶神経回路
青は核染色、緑色はDG、赤色はDGの軸索(DGからのシナプス入力)、オレンジ色はCA2を示す。実験の結果、DGが直接シナプスを介してCA2に入力していることが分かった。



